合肥細(xì)胞外基質(zhì)膠服務(wù)電話 蘇州君欣生物科技供應(yīng)
發(fā)貨地點:江蘇省蘇州市
發(fā)布時間:2025-02-11
發(fā)貨地點:江蘇省蘇州市
發(fā)布時間:2025-02-11
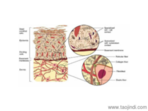
細(xì)胞外基質(zhì)與醫(yī)學(xué):惡性部位的發(fā)生、發(fā)展、侵襲和轉(zhuǎn)移常常伴有細(xì)胞外基質(zhì)(extracellular matrix,ECM)及其細(xì)胞表面受體表達(dá)的變化。正常肝細(xì)胞沒有基膜,也不表達(dá)層粘連蛋白(laminin,LN)的特異性整合素族受體α6β1;而在肝細(xì)胞(human hepatocellular carcinoma,HCC)組織中,LN和α6β1不表達(dá)水平升高,呈明顯的共分布,而且其高水平表達(dá)與肝患者的預(yù)后呈負(fù)相關(guān),合肥細(xì)胞外基質(zhì)膠服務(wù)電話,提示HCC細(xì)胞可能通過α6β1受體接受來自LN的信號,從而對肝細(xì)胞的侵襲行為起著不可忽視的作用。肝的發(fā)病過程中往往早期就出現(xiàn)門靜脈侵襲、肝內(nèi)轉(zhuǎn)移以及肝外肺臟和骨組織的轉(zhuǎn)移,肝的侵襲、轉(zhuǎn)移和術(shù)后復(fù)發(fā)是影響患者預(yù)后的主要因素。基質(zhì)金屬蛋白酶(matrix meta-lloproteinases,合肥細(xì)胞外基質(zhì)膠服務(wù)電話,MMPs)對ECM的降解是部位細(xì)胞侵襲和轉(zhuǎn)移的關(guān)鍵環(huán)節(jié)之一,多種惡性部位都伴有MMPs分泌水平和活性的增高,合肥細(xì)胞外基質(zhì)膠服務(wù)電話。而在層粘連蛋白上則停止增殖,進(jìn)行分化,融合為肌管。合肥細(xì)胞外基質(zhì)膠服務(wù)電話

細(xì)胞外基質(zhì)與醫(yī)學(xué):目前學(xué)者們一致認(rèn)為惡性部位的侵蝕、轉(zhuǎn)移是一個動態(tài)的、連續(xù)的過程。部位細(xì)胞先從原發(fā)部位脫落,侵入到細(xì)胞外基質(zhì)(extracellular ma-tric,ECM),與基底膜(basement membrane,BM)與細(xì)胞間質(zhì)中一些分子粘附,并啟動細(xì)胞合成、分泌各種降解酶類,協(xié)助部位細(xì)胞穿過ECM進(jìn)入血管,然后在某些因子等的作用下運(yùn)行并穿過血管壁外滲到繼發(fā)部位,繼續(xù)增殖、形成轉(zhuǎn)移灶。總之,脫落、粘附、降解、移動和增生貫穿于惡性部位侵蝕、轉(zhuǎn)移的全過程。ECM由BM和細(xì)胞間質(zhì)組成,為部位轉(zhuǎn)移的重要組織屏障。部位細(xì)胞通過其表面受體與ECM中的各種成分粘附后啟動或分泌蛋白降解酶類來降解基質(zhì),從而形成局部溶解區(qū),構(gòu)成了部位細(xì)胞轉(zhuǎn)移運(yùn)行通道。一般惡性程度高的部位細(xì)胞具有較強(qiáng)的蛋白水解作用,可侵蝕破壞包膜,促進(jìn)轉(zhuǎn)移。目前較為關(guān)注的酶主要是絲氨酸蛋白酶類,如纖溶酶原啟動物(plasminogen activator,PA)和金屬蛋白酶(metalproteinase,MP)類,如膠原酶IV、基質(zhì)降解酶、透明質(zhì)酸酶.。開封正規(guī)細(xì)胞外基質(zhì)膠直銷廠家不同細(xì)胞具有不同的細(xì)胞外基質(zhì),介導(dǎo)的細(xì)胞骨架組裝的狀況不同。

細(xì)胞外基質(zhì)深入研究該團(tuán)隊提出:細(xì)胞外基質(zhì)內(nèi)成分豐富,其提取方法多樣,具有指導(dǎo)細(xì)胞、傳遞信號、診治疾病及再生修復(fù)等眾多功能。細(xì)胞外基質(zhì)作為一種精密有序的網(wǎng)絡(luò)結(jié)構(gòu)在組織再生與修復(fù)中具有獨(dú)特的優(yōu)勢,它能提供較接近于體內(nèi)細(xì)胞生長的微環(huán)境,加之其富含的各種活性分子可為細(xì)胞活動提供基礎(chǔ),具有較好的應(yīng)用前景。細(xì)胞外基質(zhì)在組織再生中以多種形式存在,其優(yōu)點在于具有良好的生物降解性和較低的免疫原性,并能在一定程度上改善炎癥反應(yīng),可有效支持組織再生并指導(dǎo)組織重建。隨著人們對細(xì)胞外基質(zhì)的不斷深入研究,其在組織工程中的應(yīng)用將會獲得更好的發(fā)展,也將為再生醫(yī)學(xué)及組織工程的未來帶來新興技術(shù)及方法,但對于細(xì)胞外基質(zhì)在組織修復(fù)應(yīng)用中面臨的問題,還需要更深入的研究和探討。
涉及纖連蛋白基質(zhì)組裝的信號傳導(dǎo)途徑:原纖維形成檢測(貨號:FNR01,F(xiàn)NR02,F(xiàn)NR03)與其他可以在生理條件下自動聚合的ECM組分不同,F(xiàn)ibronectin的組裝是一種依賴細(xì)胞的過程。了解FN組裝所涉及的機(jī)制以及這些與細(xì)胞,纖維化和免疫反應(yīng)之間的相互作用可能揭示了調(diào)控異常組織修復(fù)過程的療法的未來發(fā)展目標(biāo)。同樣,組織工程很大程度上依賴于控制細(xì)胞外基質(zhì)形成的速率和模式的能力。Cytoskeleton熒光標(biāo)記的Fibronectin可用于檢測原纖維形成。熒光纖連蛋白(FNR01 和FNR02該方法涉及通過摻入熒光纖連蛋白來對纖維絲形成進(jìn)行熒光示蹤(17),通過向細(xì)胞培養(yǎng)基中加入TRITC標(biāo)記的Fibronectin(FNR01)或HiLyte488標(biāo)記的Fibronectin(FNR02),可以觀察到可溶性Fibronectin向細(xì)胞表面不溶纖維絲的轉(zhuǎn)化,摻入的纖連蛋白的水平可以通過熒光顯微鏡觀察和定量(18)。在層粘連蛋白上則停止增殖,進(jìn)行分化,融合為肌管。
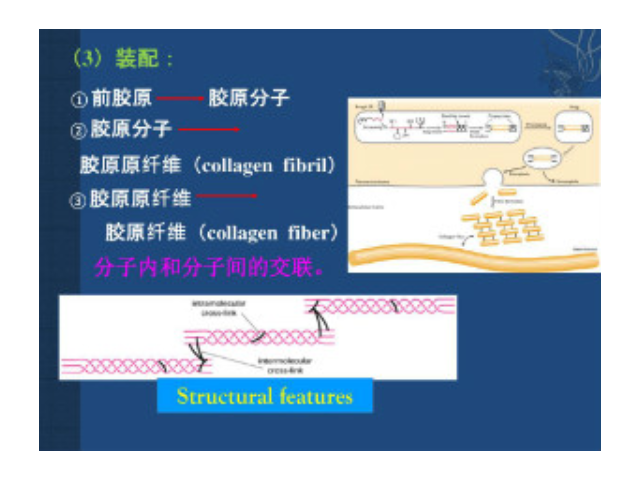
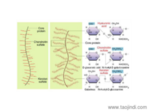
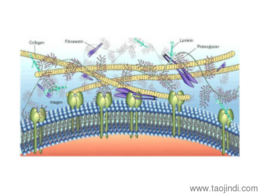
根據(jù)膠原的結(jié)構(gòu)和功能可將其分為:纖維性膠原(fibril forming collagen) 這是較經(jīng)典的膠原,如Ⅰ、Ⅲ、Ⅴ和Ⅺ型膠原。其肽鏈長達(dá)1000個氨基酸,是結(jié)締組織中含量較豐富的膠原。前膠原三螺旋的端肽被切除后縱向平行排列,其中每個膠原分子縱向稍偏移,相鄰的肽鏈形成共價鍵交聯(lián)從而形成微纖維。一般需經(jīng)前膠原肽酶(procollagen propeptidase)將羧基端肽去除后才能形成膠原纖維,但是部分膠原可以帶有氨基端肽而存在于膠原纖維的表面,以阻止膠原纖維繼續(xù)增粗,從而繼續(xù)起到調(diào)節(jié)膠原纖維直徑的作用。主要是一些多糖和蛋白, 或蛋白聚糖。合肥細(xì)胞外基質(zhì)膠服務(wù)電話
蛋白聚糖,由蛋白質(zhì)和多糖共價形成,具有高度親水性。合肥細(xì)胞外基質(zhì)膠服務(wù)電話
細(xì)胞外基質(zhì)偶聯(lián)調(diào)控細(xì)胞單層的受力反應(yīng):細(xì)胞和基底之間的耦合(通過粘附分子)屬于高度非線性耦合;細(xì)胞外牽引力和細(xì)胞間張力與細(xì)胞單層尺寸相關(guān),也與基質(zhì)剛度有關(guān),提示細(xì)胞內(nèi)、外分子之間存在相互信息交流;在細(xì)胞單層邊緣,粘著斑通過與細(xì)胞外基質(zhì)的相互作用產(chǎn)生了強(qiáng)大牽引力,而在細(xì)胞單層的中央,細(xì)胞應(yīng)激水平上升的同時遭受到較低牽引力。這些結(jié)果將有助于解釋局部環(huán)境如何影響細(xì)胞決策,也將促進(jìn)上皮組織中諸如形態(tài)發(fā)生或集體遷移等更為復(fù)雜問題的研究。綜上,該模型為多用途計算框架奠定了堅實的基礎(chǔ),可用來揭示多細(xì)胞上皮形態(tài)發(fā)生和疾病的分子起源。合肥細(xì)胞外基質(zhì)膠服務(wù)電話