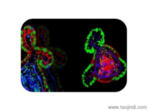
細胞外基質的成分:膠原(collagen)膠原是動物體內含量較豐富的蛋白質,約占人體蛋白質總量的30%以上。它遍布于體內各種部位和組織,是細胞外基質中的框架結構,可由成纖維細胞、軟骨細胞、成骨細胞及某些上皮細胞合成并分泌到細胞外。成纖維細胞周圍的膠原纖維目前已發現的膠原至少有19種(,由不同的結構基因編碼,無錫正規細胞外基質膠,具有不同的化學結構及免疫學特性。Ⅰ、Ⅱ、Ⅲ、Ⅴ及Ⅺ型膠原為有橫紋的纖維形膠原,無錫正規細胞外基質膠。各型膠原都是由三條相同或不同的肽鏈形成三股螺旋,含有三種結構:螺旋區,無錫正規細胞外基質膠,非螺旋區及球形結構域。其中Ⅰ型膠原的結構較為典型。彈性蛋白由二種類型短肽段交替排列構成。無錫正規細胞外基質膠

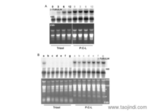
骨膜來源的細胞外基質水凝膠通過早期免疫調節及增強血管和骨生成促進骨修復:骨愈合包括早期炎癥免疫調節、血管生成、成骨分化和生物礦化等過程,干預其中的任一過程都可能阻礙骨修復。在復雜和嚴重的骨損傷部位,大量的促炎因子的存在會誘發促炎反應。長期的促炎反應會阻礙巨噬細胞從M1到M2的轉變導致骨再生延遲。因此,在骨損傷早期通過調控M1向M2的轉變來適時終止促炎反應是骨愈合成功的前提。近日,浙江大學醫學院范順武和林賢豐教授課題組制備了一種骨膜來源的細胞外基質(PEM)水凝膠,并評價了它們在骨修復過程中不同時期的調節作用。無錫正規細胞外基質膠破壞了腎小球的組織結構,損傷了腎小球的功能,較終導致腎小球硬化的形成。
細胞外基質成分居然能調節葡萄糖代謝過程:確認ECM組分透明質酸與新陳代謝之間的機制聯系,用透明質酸酶處理細胞和異種移植物引發糖酵解的強烈增加。這主要通過快速受體酪氨酸激酶介導的mRNA衰變因子ZFP36的誘導來實現,其靶向TXNIP轉錄物以降解。因為TXNIP促進葡萄糖轉運蛋白GLUT1的內化,其急劇下降使質膜上的GLUT1富集。在功能上,需要通過透明質酸酶誘導糖酵解,同時加速細胞遷移。 ECM重塑和代謝之間的這種相互關聯在動態組織狀態中表現出來,包括部位發生和胚胎發生。研究人員提出ECM重塑作為急性細胞 - 外在代謝調節的附加節點,可以直接讀出周圍組織的結構狀態并相應地增強細胞行為。
免疫系統和細胞外基質之間的串擾:ECM是三維網狀,支持細胞,調節重要的細胞過程:增殖,粘附,遷移,細胞分化和炎癥。在對損傷的反應中,先發生的事件包括免疫系統的啟動和基質金屬蛋白酶(MMPs)的上調。細胞對損傷信號的反應進程和較終結果在一定程度上受創床中存在的特定MMP及其活性持續時間的控制。克制巨噬細胞募集到損傷部位已被證明可以克制再生;然而,在體內對ECM重塑的影響研究較少。細胞外基質和免疫系統之間的這種相互作用在再生物種中是如何工作的尚不清楚。刺胞動物免疫系統的主要調節因子是蛋白酶、絲氨酸蛋白酶克制劑、克菌蛋白和補體系統。免疫的原始機制是克菌肽(AMPs),在水螅體再生過程中,一些被歸類為AMPs的基因被上調。蛋白聚糖,由蛋白質和多糖共價形成,具有高度親水性。

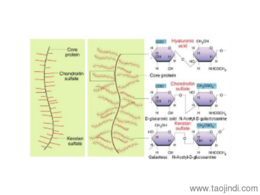
細胞外基質氨基聚糖與蛋白聚氨基聚糖(glycosaminoglycan,GAG) GAG是由重復二糖單位構成的無分枝長鏈多糖。其二糖單位通常由氨基已糖(氨基葡萄糖或氨基半乳糖)和糖醛酸組成,但硫酸角質素中糖醛酸由半乳糖代替。氨基聚糖依組成糖基、連接方式、硫酸化程度及位置的不同可分為六種,即:透明質酸、硫酸軟骨素、硫酸皮膚素、硫酸乙酰肝素、肝素、硫酸角質素。透明質酸(hyaluronic acid,HA)是較少不發生硫酸化的氨基聚糖,其糖鏈特別長。氨基聚糖一般由不到300個單糖基組成,而HA可含10萬個糖基。在溶液中HA分子呈無規則卷曲狀態。如果強行伸長,其分子長度可達20μm。HA整個分子全部由葡萄糖醛酸及乙酰氨基葡萄糖二糖單位重復排列構成。由于HA分子表面有大量帶負電荷的親水性基團,可結合大量水分子,因而即使濃度很低也能形成粘稠的膠體,占據很大的空間,產生膨壓。成纖維細胞在纖粘連蛋白基質上增殖加快。無錫正規細胞外基質膠
細胞外基質不靜態的發揮支持、連接、保水、保護等物理作用。無錫正規細胞外基質膠
細胞外基質偶聯調控細胞單層的受力反應:細胞和基底之間的耦合(通過粘附分子)屬于高度非線性耦合;細胞外牽引力和細胞間張力與細胞單層尺寸相關,也與基質剛度有關,提示細胞內、外分子之間存在相互信息交流;在細胞單層邊緣,粘著斑通過與細胞外基質的相互作用產生了強大牽引力,而在細胞單層的中央,細胞應激水平上升的同時遭受到較低牽引力。這些結果將有助于解釋局部環境如何影響細胞決策,也將促進上皮組織中諸如形態發生或集體遷移等更為復雜問題的研究。綜上,該模型為多用途計算框架奠定了堅實的基礎,可用來揭示多細胞上皮形態發生和疾病的分子起源。無錫正規細胞外基質膠