江蘇疫苗熱原檢測(cè)規(guī)范
來(lái)源:
發(fā)布時(shí)間:2025-11-12
在 MAT 熱原檢測(cè)中,單核細(xì)胞系與 PBMC(外周血單個(gè)核細(xì)胞)的檢測(cè)結(jié)果穩(wěn)定性差異明顯。實(shí)驗(yàn)結(jié)果數(shù)據(jù)顯示,單核細(xì)胞系的標(biāo)曲 R2 達(dá) 1.000,各濃度點(diǎn) CV 值極低(如 ST1 為 0.92%、ST2 為 1.5%),相對(duì)偏差均在 5% 以內(nèi);而 PBMC 的標(biāo)曲 R2 為 0.997,部分濃度點(diǎn) CV 值超 20%(如 ST2 為 39.6%、ST6 為 45.5%),相對(duì)偏差高達(dá) 171.43%。這種差異源于兩者對(duì)熱原反應(yīng)的一致性—單核細(xì)胞系能穩(wěn)定釋放 IL-6,PBMC 則因供體差異導(dǎo)致 IL-6 釋放水平波動(dòng),直接影響熱原檢測(cè)結(jié)果的重復(fù)性,單核細(xì)胞系更適配準(zhǔn)確的熱原定量需求。
鱟試驗(yàn)法(LAL)法進(jìn)行熱原檢測(cè)靈敏度高、操作方便,但只針對(duì)內(nèi)毒素,易受干擾且有 LER 現(xiàn)象。江蘇疫苗熱原檢測(cè)規(guī)范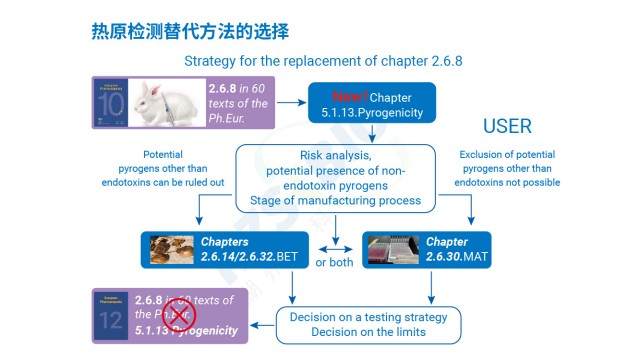
熱原是指微量即可引發(fā)恒溫動(dòng)物體溫異常升高的物質(zhì),分為內(nèi)源性(如細(xì)胞因子)與外源性兩類(lèi),外源性熱原又涵蓋微生物來(lái)源(革蘭氏陰性菌脂多糖 LPS、革蘭氏陽(yáng)性菌脂磷壁酸 LTA、病毒、真菌等)與非微生物來(lái)源(灰塵、橡膠降解產(chǎn)物等)。傳統(tǒng)細(xì)菌內(nèi)毒素檢查法(BET)只能檢測(cè)革蘭氏陰性菌的 LPS,無(wú)法覆蓋非內(nèi)毒素?zé)嵩鴨魏思?xì)胞活化試驗(yàn)(MAT)可彌補(bǔ)這一缺陷。其原理是:熱原通過(guò)活化單核細(xì)胞表面的 Toll 樣受體(TLR,如 TLR4 識(shí)別 LPS、TLR2/TLR6 識(shí)別 LTA),啟動(dòng)先天免疫反應(yīng),促使細(xì)胞釋放 IL-6、TNF-α 等促炎細(xì)胞因子;隨后采用 ELISA 法檢測(cè) IL-6 濃度,結(jié)合 LPS 標(biāo)準(zhǔn)曲線推算樣品中總熱原含量,實(shí)現(xiàn)對(duì)內(nèi)毒素與非內(nèi)毒素?zé)嵩耐綑z測(cè),契合《中國(guó)藥典》9301 指導(dǎo)原則中 全場(chǎng)景防控?zé)嵩L(fēng)險(xiǎn)”的要求。
江蘇合規(guī)性熱原檢測(cè)流程單核細(xì)胞活化試驗(yàn)(MAT)將熱原檢測(cè)從經(jīng)驗(yàn)性觀察,推進(jìn)至受體-配體相互作用的分子本質(zhì)。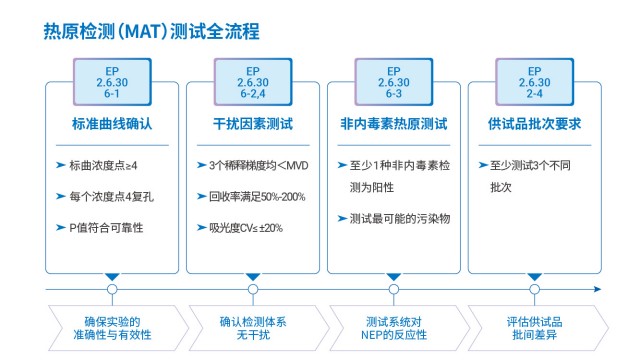
傳統(tǒng)熱原檢測(cè)方法的局限性十分明顯:鱟試驗(yàn)法只能特異性識(shí)別細(xì)菌內(nèi)毒素,對(duì)非內(nèi)毒素?zé)嵩耆珶o(wú)響應(yīng);家兔熱原試驗(yàn)雖能篩查全類(lèi)型熱原,但靈敏度低(檢測(cè)限≥5EU/kg),無(wú)法檢出微量非內(nèi)毒素?zé)嵩覠o(wú)法區(qū)分熱原類(lèi)型,難以追溯污染源頭;單核細(xì)胞活化反應(yīng)測(cè)定(MAT)的出現(xiàn)有效解決了上述難題,其關(guān)鍵優(yōu)勢(shì)在于:基于人源單核細(xì)胞的免疫應(yīng)答機(jī)制,可同時(shí)識(shí)別所有具備生物活性的熱原(包括內(nèi)毒素與非內(nèi)毒素),且檢測(cè)靈敏度高(對(duì)病毒熱原檢測(cè)限低至pg級(jí));檢測(cè)周期短(24-48小時(shí)),可滿足產(chǎn)品快速放行需求;能通過(guò)細(xì)胞因子濃度定量評(píng)估熱原活性,避免“無(wú)活性熱原”的誤判。
在 MAT 法熱原檢測(cè)中,PBMC(外周血單核細(xì)胞)與單核細(xì)胞系各有優(yōu)劣,單核細(xì)胞系更適合標(biāo)準(zhǔn)化檢測(cè)。PBMC 的優(yōu)勢(shì)在于免疫細(xì)胞成分豐富(含單核細(xì)胞、淋巴細(xì)胞等),對(duì)熱原反應(yīng)敏感,靈敏度相對(duì)較高;但局限同樣明顯 ——PBMC 需從不同供體獲取,供體免疫狀態(tài)差異會(huì)導(dǎo)致檢測(cè)結(jié)果不穩(wěn)定,且無(wú)法長(zhǎng)期保存,難以建立標(biāo)準(zhǔn)化方法學(xué)。單核細(xì)胞系(如 HL-60、MM6、THP1)則克服了 PBMC 的局限:細(xì)胞來(lái)源穩(wěn)定(可批量培養(yǎng)),TLR 受體表達(dá)覆蓋主要亞型(如 HL-60 表達(dá) TLR1-TLR9),對(duì)熱原反應(yīng)重復(fù)性好,更適合商業(yè)化試劑盒與法規(guī)檢測(cè)。不同單核細(xì)胞系性能也有差異:MM6/IL-6 法檢測(cè)限約 0.05EU/mL,THP1/TNF-α 法因 TNF-α 為一級(jí)免疫效應(yīng)物檢測(cè)限更低,但 TNF-α 穩(wěn)定性差;HL-60/IL-6 法檢測(cè)限與穩(wěn)定性均優(yōu)于前兩者,成為主流選擇。湖州申科生物MAT試劑盒選用 HL-60 細(xì)胞系,正是基于其優(yōu)異的穩(wěn)定性與熱原響應(yīng)適配多場(chǎng)景,確保不同批次檢測(cè)結(jié)果一致。
當(dāng)熱原侵入人體循環(huán)系統(tǒng),單核細(xì)胞表面的TLR即刻化身分子雷達(dá),準(zhǔn)確捕獲LPS、LTA等外源致熱物。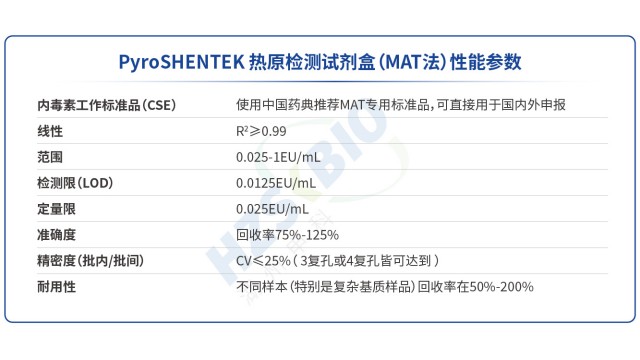
傳統(tǒng)細(xì)菌內(nèi)毒素檢查法(BET)只能檢測(cè)革蘭氏陰性菌的 LPS,無(wú)法識(shí)別革蘭氏陽(yáng)性菌 LTA、真菌酵母多糖、病毒鞭毛蛋白等非內(nèi)毒素?zé)嵩嬖诼z風(fēng)險(xiǎn);同時(shí),部分樣品(如脂質(zhì)體、表面活性劑制劑)會(huì)因內(nèi)毒素吸附導(dǎo)致低內(nèi)毒素回收(LER),BET法難以準(zhǔn)確定量。MAT 法通過(guò)單核細(xì)胞表面的多種 TLR 受體(TLR1-TLR10),可識(shí)別不同類(lèi)型熱原:TLR4 識(shí)別 LPS、TLR2/TLR6 識(shí)別 LTA 與酵母多糖、TLR5 識(shí)別鞭毛蛋白、TLR3 識(shí)別病毒 dsRNA 等,實(shí)現(xiàn) “全熱原覆蓋”。湖州申科生物熱原檢測(cè)試劑盒(MAT法)的驗(yàn)證數(shù)據(jù)顯示,其對(duì)不同濃度非內(nèi)毒素?zé)嵩许憫?yīng):如 0.1-100μg/mL 鞭毛蛋白可檢測(cè)到 0.005-0.035EU/mL熱原活性,0.1-10μg/mL LTA 對(duì)應(yīng) 0.1-0.7EU/mL 熱原活性,1-100μg/mL 雷西莫特(TLR7/8 配體)對(duì)應(yīng) 0.5-3.0EU/mL 熱原活性。此外,MAT法檢測(cè)的是熱原的生物活性(而非單純 LPS 含量),可避免 LER 導(dǎo)致的假陰性,為CGT等高風(fēng)險(xiǎn)產(chǎn)品提供更有保障的熱原檢測(cè)方案。
湖州申科熱原檢測(cè)試劑盒聯(lián)合了國(guó)內(nèi)相關(guān)機(jī)構(gòu)室間驗(yàn)證,與傳統(tǒng)RPT法結(jié)果高度一致,符合法規(guī)要求。江蘇合規(guī)性熱原檢測(cè)流程生物制品的高蛋白、螯合劑基質(zhì)易對(duì)鱟試驗(yàn)產(chǎn)生抑制,rCR與MAT聯(lián)合策略可消除干擾并控制熱原。江蘇疫苗熱原檢測(cè)規(guī)范
PyroSHENTEK®熱原檢測(cè)試劑盒以 “簡(jiǎn)化流程、提升效率” 為設(shè)計(jì)理念,采用即用型細(xì)胞,凍存細(xì)胞無(wú)需離心復(fù)蘇培養(yǎng),復(fù)融后可直接與供試品混合共孵育,大幅節(jié)省傳統(tǒng)細(xì)胞預(yù)處理(如調(diào)整細(xì)胞狀態(tài)、鋪板培養(yǎng))的時(shí)間成本。實(shí)驗(yàn)流程清晰可控:只需按要求制備待測(cè)供試品與內(nèi)毒素工作標(biāo)準(zhǔn)品,各取對(duì)應(yīng)體積加入細(xì)胞懸液(每孔加樣量明確),經(jīng) 37℃、5% CO?孵育 24 小時(shí)后,離心收集上清并通過(guò) ELISA 法檢測(cè) IL-6 含量,全程可在 24 小時(shí)內(nèi)獲得穩(wěn)定可靠的檢測(cè)結(jié)果。這種便捷性尤其適配實(shí)驗(yàn)室高通量檢測(cè)需求,減少人員操作步驟的同時(shí),降低了因復(fù)雜操作引入的誤差風(fēng)險(xiǎn),兼顧效率與準(zhǔn)確性。
江蘇疫苗熱原檢測(cè)規(guī)范
相關(guān)新聞
- 上海高效熱原檢測(cè)MAT試劑盒 2025-12-19
- 北京高效內(nèi)毒素檢測(cè)商業(yè)化試劑盒 2025-12-19
- 北京通用型宿主細(xì)胞蛋白(HCP)殘留檢測(cè)抗體制備 2025-12-19
- 江蘇生物制品內(nèi)毒素檢測(cè)重組級(jí)聯(lián)試劑(rCR) 2025-12-19
- 北京通用型宿主細(xì)胞蛋白(HCP)殘留檢測(cè) 2025-12-19
- 浙江定制化宿主細(xì)胞蛋白(HCP)殘留檢測(cè)抗體覆蓋率驗(yàn)證 2025-12-19
- SV40LTA&E1A宿主細(xì)胞殘留DNA檢測(cè) 2025-12-19
- 江蘇細(xì)菌內(nèi)毒素檢測(cè)結(jié)果判定 2025-12-19
- 上海通用型宿主細(xì)胞蛋白(HCP)殘留檢測(cè)方法學(xué)驗(yàn)證 2025-12-19
- 北京ELISA法宿主細(xì)胞蛋白(HCP)殘留檢測(cè)抗體制備 2025-12-19
推薦新聞
- 佛山1470nm國(guó)產(chǎn)半導(dǎo)體激光治療儀廠家電話 2025-12-19
- 吳江區(qū)服務(wù)軟霧吸入器加盟連鎖店 2025-12-19
- 松江區(qū)定制吻合器工廠直銷(xiāo) 2025-12-19
- 深圳醫(yī)院智慧管理系統(tǒng)多少錢(qián)一套 2025-12-19
- 山西疫苗佐劑QS-21 2025-12-19
- 姑蘇區(qū)常見(jiàn)一次性注射器資費(fèi) 2025-12-19
- 武漢本地活細(xì)胞成像分析系統(tǒng)直銷(xiāo)價(jià)格 2025-12-19
- 附近哪里有助腿器誠(chéng)信合作 2025-12-19
- 銀杏葉片工廠 2025-12-19
- 珠海有實(shí)力的nmn定制廠家 2025-12-19